Sistema Imunológico Humano: uma revisão atualizada
Prof Ulysses Fagundes Neto
Introdução
Tendo em vista a presente pandemia do Sars-cov-2 que nos assola há 9 meses, e que se mostra resistente com altos números de morbidade e letalidade, nas mais distintas regiões do Brasil, e, considerando-se o papel primordial do nosso sistema imunológico no seu combate, vale a pena revisarmos a interação do agente agressor com nosso sistema de defesa. Tem sido amplamente divulgado que a resposta do nosso organismo ao Sars-cov-2 tem profundas implicações no sistema imunológico, e, portanto, uma atualização básica do sistema imunológico humano, torna-se essencial, ainda que fugindo ao escopo primordial deste meio de comunicação, isto é, a Gastroenterologia e a Nutrição.
O sistema imunológico trata-se de uma complexa rede de diferentes elementos que interagem entre si. Quando um agente patogênico invade nosso organismo, imediatamente é desencadeada uma resposta imunológica inata. Enquanto muitas infecções podem ser devidamente resolvidas por meio do sistema imunológico inato, os seres humanos também possuem um sistema adicional de defesa, a imunidade adaptativa, a qual se adapta para nos proteger contra invasores específicos. A seguir, passaremos a abordar ambos os componentes do nosso sistema imunológico.
Imunidade Inata
A primeira linha do mecanismo de defesa, contra microrganismos estranhos aos seres humanos, é mediada pelo sistema imunológico inato. As células chave envolvidas no processo inflamatório incluem neutrófilos, macrófagos, células dendríticas e células natural killers (NK). Estas células utilizam mecanismos similares de reconhecimento de proteínas celulares para identificar células estranhas e removê-las do nosso organismo (Figura 1).
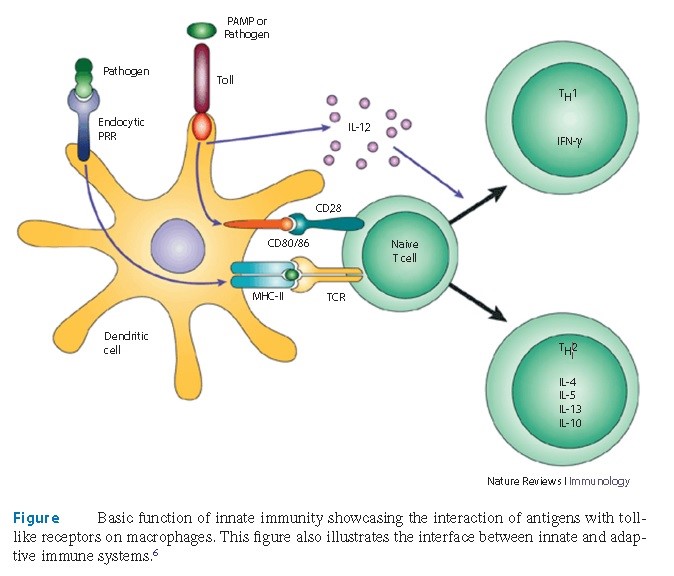
Figura 1
Proteínas de reconhecimento padrão (PRP) representam os componentes essenciais da imunidade inata. Neutrófilos e macrófagos possuem receptores proteicos nas suas membranas celulares que reconhecem determinados padrões de células estranhas, e não somente as reconhecem como estranhas, como também as diferenciam das células hospedeiras durante a sua eliminação. Como exemplos de padrões moleculares associados aos patógenos estão as endotoxinas bacterianas das bactérias Gram negativas, peptídeoglicans, ácido lipoteicoico e a dupla banda de RNA viral (Figura 2).
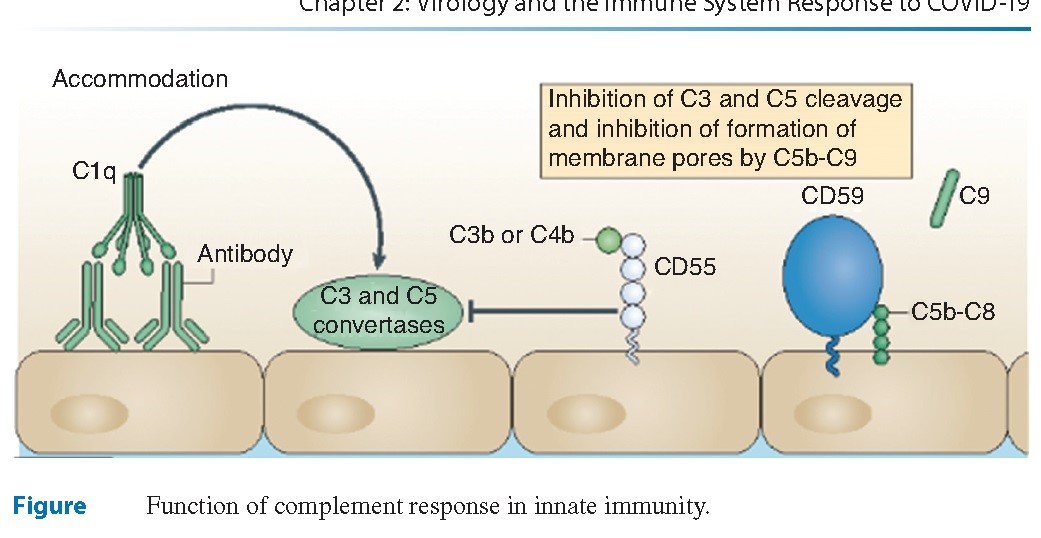
Figura 2
Um dos exemplos mais importantes e bem estudados dos PRPs é o sistema complemento, que se constitui em uma série de peptídeos circulantes que desempenham um papel crucial na imunidade inata do hospedeiro. Um componente fundamental desse sistema, a proteína C1, reconhece anticorpos ligados às proteínas microbianas e inicia um evento em cadeia, conhecido como a cascata do complemento. O resultado desta atividade é a produção do C3B, que provoca a quimiotaxia e a fagocitose da bactéria que culmina na formação de um complexo de ataque à membrana.
Os principais componentes do sistema imunológico inato são: neutrófilos, macrófagos e células NK. Os neutrófilos são direcionados para os locais da inflamação ou infecção, pelas citocinas, tais como as proteínas IL-8 e C3B. Os macrófagos também apresentam uma função similar, porém, eles respondem de uma forma um pouco mais lenta do que os neutrófilos. As células NK contém receptores proteicos similares, os quais são utilizados para diferenciar as células dos hospedeiros e das infectantes. Uma vez feito o reconhecimento, as células NK induzem a apoptose. As células NK são especialmente importantes na proteção contra células virais e tumorais. Outros componentes importantes do sistema imunológico inato são os eosinófilos, basófilos e mastócitos.
Uma característica importante do sistema imunológico inato é que ele não dispõe de células de memória ou respostas aprendidas contra antígenos específicos. O sistema imunológico inato apresenta uma característica genérica e inespecífica à inflamação. Diferentemente do sistema imunológico adaptativo que possui uma sintonia fina e se auto modifica para combater uma infecção específica, durante um longo período, o sistema imunológico inato é desenhado para apresentar respostas imediatas. É importante ressaltar que o sistema imunológico inato representa o processo preliminar necessário para gerar o sistema imunológico adaptativo.
Imunidade Adaptativa
O sistema imunológico adaptativo, diferentemente do inato, trata-se de um mecanismo mais refinado e programado da defesa do hospedeiro. A resposta elaborada da imunidade adaptativa ocorre a longo prazo em comparação com a resposta instantânea e generalizada do sistema imunológico inato. Existem dois tipos de respostas imunológicas adaptativas, a saber: imunidade humoral e imunidade mediada por células, que a seguir serão descritas.
Imunidade Humoral
A imunidade humoral (IH) é organizada pelas células B que secretam anticorpos para a proteção do organismo contra microrganismos estranhos de forma imediata, e, também de longo prazo. O evento crucial envolve receptores nas células B que se ligam a um antígeno. Esta ligação, provoca a ativação e a diferenciação em células mais especializadas. As células B consistem em um receptor que é constituído por cadeias de imunoglobulinas leves e pesadas, e um correceptor de células B, o qual é necessário para uma ligação eficaz ao antígeno. A ligação de antígenos a múltiplos receptores das células B, provoca uma serie de fosforilações intracelulares via a rota IP3 e diacil glicerol, sinalização intracelular, a qual resulta na translocação de fatores de transcrição para o núcleo e ativação das células B.
Desta forma as células B podem se ligar a antígenos timo-dependentes e timo-independentes. Os clássicos exemplos de antígenos timo-independentes são os polissacarídeos nos organismos capsulares, tais como, H influenza e S pneumonia, que acarreta a organização da nossa resposta sorológica de anticorpos. Este mecanismo é também a base para as vacinações dos polissacarídeos não conjugados. Essas cápsulas contêm açúcar e não são apropriados para uma resposta mais robusta às vacinas mediadas por células T. Entretanto, as células B são as candidatas ideais para responder a estes patógenos estranhos.
O antígeno timo-dependente provoca uma especialização das células B que as fazem assumir papéis mais específicos. A ligação destes antígenos ativa as células T-helper. Estas células T em sequência ativam as células B para secretarem tipos específicos de anticorpos. Por exemplo, nos centros germinais do tecido linfoide secundário, a ligação de células B carream antígeno para a célula Th2 e seu respectivo correceptor, provoca a liberação de IL-4 e IL-5, as quais sinalizam para as células B para secretarem IgE. Anticorpos IgE tem o poder de proteger contra infestações helmínticas e enfermidades atópicas.
No interior dos centros germinativos do tecido linfoide, cada célula B é desafiada para a realização de tarefas essenciais para sobreviver e proliferar. Estas tarefas incluem competir com outras células B pela ligação com antígenos, o processamento de antígenos, e a apresentação deles para as células T–helper, recebendo sinalizações estimulantes de correceptores, e, então finalmente, se especializando. Células B altamente especializadas e diferenciadas podem também circular no plasma como célula B de memória, onde elas têm a lembrança dos antígenos com os quais interagiram no passado e, desta forma, passar à produção de anticorpos.
As células B também desempenham um papel crucial na imunidade inata, e, como anteriormente abordado, o sistema complemento é um componente chave da imunidade inata. O antígeno ligado a regiões Fab dos anticorpos IgG apresentam uma ligação cruzada com as regiões Fc dos anticorpos. Essencialmente o anticorpo funciona como uma ponte para o complemento e o antígeno. Uma vez que a ponte é construída, o complemento pode solicitar o recrutamento de neutrófilos e posteriormente ocorre a fagocitose do microrganismo.
Imunidade Celular
A imunidade celular é um vigoroso sistema de mecanismos de defesa do hospedeiro, e que foi desenhada para combater microrganismos intracelulares, tais como vírus e micobactérias, bem como células tumorais. Este mesmo sistema também é “o culpado” por estar envolvido em muitas condições autoimunes. As células T são os condutores primários da imunidade mediada por células. Embora existam diversos tipos de células T, as células CD4 e CD8 expressas pelos linfócitos T citotóxicos são importantes na avaliação da imunidade dos pacientes (Figura 3).
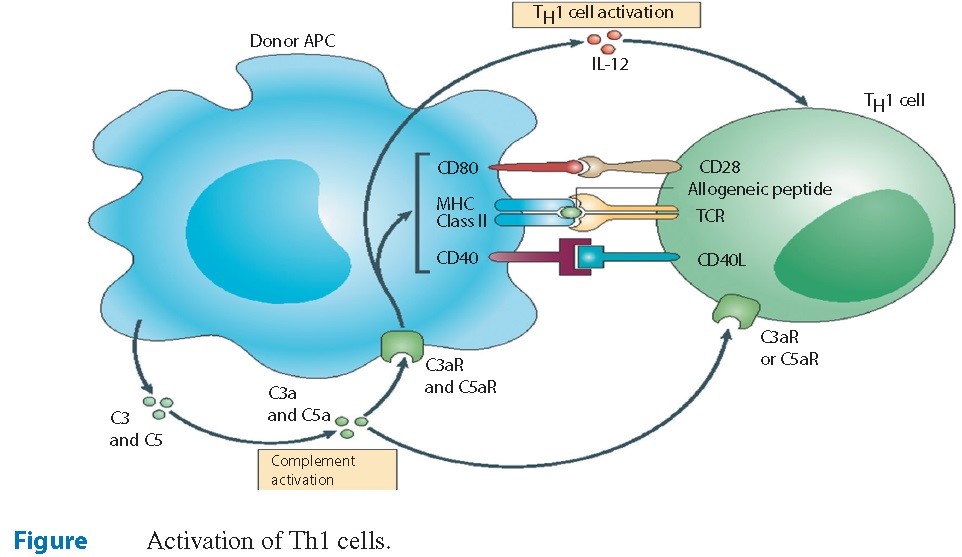
Figura 3
As células CD8+ são células citotóxicas e que possuem mecanismos citotóxicos contra células infectadas, com as quais após ocorrer a ligação, causam uma fusão e liberação de grânulos que acarretam dano celular. As células CD8+ também expressam ligações Fas que se ligam a receptores CD-95 das células infectadas. Estas ligações acarretam a apoptose. As células CD8+ têm capacidade própria de eliminar células infectadas por vírus e células tumorais por meio de apoptose ou do envolvimento de células NK. As células CD8+ necessitam ser ativadas pela via de apresentação de antígeno através das células MHC-1. Considerando-se que todas as células nucleadas expressam células MHC-1, CD8+ são ideais para a eliminação de células que podem ser infectadas por vírus, posto que quaisquer células podem apresentar antígenos para as células CD8+. Entretanto, antes que as células CD8+ possam adquirir essa capacidade, elas necessitam ser ativadas no tecido linfoide com o auxílio das células CD4+ e das células T apresentadoras de antígenos.
As células CD4+ são as células T-helper e são ativadas nos linfonódios quando as células dendríticas capturam antígenos estranhos e os apresentam às células T. A ligação das células T receptoras com essas células apresentadoras de antígenos devem ser acompanhadas com um sinal de ligação com as células CD-40L sobre as células CD+4 para que o efeito esperado venha a ocorrer.
A interação das células CD4+ com o antígeno determina o destino destas células T. Citocinas e mensageiros celulares conduzem essa diferenciação. Por meio de eventos na imunidade inata, IL-12 e interferon Gama são produzidos, os quais induzem a conversão das células CD+4 em células Th1. Estas células, por sua vez, secretam uma maior quantidade de IL-12 e interferon gama, as quais promovem um intercâmbio de classe das células B que passam a secretar IgG e, também, ativam os macrófagos. Os macrófagos são ativados pela liberação de IL-12 e interferon gama desde as células Th-1. Em outras palavras, estes mecanismos tornam-se um ciclo auto regulado. Células CD+4 virgens se convertem em células Th-1 pela ação de determinadas citocinas. Células Th-1 passam a secretar citocinas de forma autônoma, as quais se preparam para uma resposta imunológica robusta, que resultam em uma condição pró-inflamatória que desempenha um papel essencial na erradicação do vírus.
A imunidade mediada por células também demonstra possuir o fenômeno de células de memória. Interações com antígenos induzem alterações nas superfícies moleculares e intracelulares que permitem às células T adquirir uma resposta mais rápida e especializada quando exposta ao mesmo antígeno. Em outras palavras, as células T têm lembrança dos antígenos que elas combateram e sabem exatamente como derrotá-los caso se apresentem novamente. Este mecanismo forma a base da imunidade perene, e, é o racional que está por trás de como as vacinas funcionam.
Conclusões
Como se pode depreender da leitura deste artigo, possuímos um robusto e diversificado sistema imunológico de defesa para garantir a proteção do nosso organismo. Apesar desta imensa rede de defesa o covid-19 tem conseguido vencer esta fantástica barreira e causar um enorme desastre sanitário com vários milhões de mortes em todo o globo terrestre, até o presente momento. Enquanto uma vacinação segura e disponível em larga escala não se apresenta, resta-nos prevenir a infecção por meios físicos, ou seja, distanciamento social, uso de máscaras e álcool nas mãos, até que o imunizante possa mitigar esta pandemia, que tanto mal tem causado à população mundial